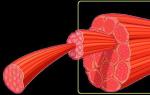
Sileää lihaskudosta säätelee vain hermosto. Sileä lihas
Kudos on kokoelma samankaltaisia soluja, joita yhdistävät yhteiset toiminnot. Lähes kaikki koostuvat erilaisista kankaista.
Luokittelu
Eläinten ja ihmisten kehossa on seuraavan tyyppisiä kudoksia:
- epiteeli;
- hermostunut;
- yhdistäminen;
- lihaksikas.
Nämä ryhmät yhdistävät useita lajikkeita. Siten sidekudos voi olla rasvaista, rustoista tai luusta. Tämä sisältää myös veren ja imunesteen. Epiteelikudos on monikerroksinen ja yksikerroksinen, solujen rakenteesta riippuen voidaan erottaa myös litteä, kuutiomainen, pylväsmäinen epiteeli jne. Hermostokudosta on vain yhtä tyyppiä. Ja puhumme siitä yksityiskohtaisemmin tässä artikkelissa.
Lihaskudoksen tyypit
Kaikkien eläinten kehossa sitä on kolme tyyppiä:
- poikkijuovaiset lihakset;
- sydämen lihaskudosta.

Sileän lihaskudoksen toiminnot eroavat poikkijuovaisen ja sydänkudoksen toiminnoista, joten sen rakenne on erilainen. Katsotaanpa tarkemmin kunkin lihastyypin rakennetta.
Lihaskudoksen yleiset ominaisuudet
Koska kaikki kolme lajia kuuluvat samaan tyyppiin, niillä on paljon yhteistä.
Lihaskudossoluja kutsutaan myosyyteiksi tai kuiduiksi. Kangastyypistä riippuen niillä voi olla erilainen rakenne.
Toinen yhteinen piirre kaikentyyppisille lihaksille on, että ne pystyvät supistumaan, mutta tämä prosessi tapahtuu yksilöllisesti eri lajeissa.
Myosyyttien ominaisuudet
Sileät lihassolut, kuten poikkijuovainen ja sydänkudos, ovat muodoltaan pitkänomaisia. Lisäksi heillä on erityisiä organelleja, joita kutsutaan myofibrilleiksi tai myofilamenteiksi. Ne sisältävät (aktiinia, myosiinia). Ne ovat välttämättömiä lihasten liikkeen varmistamiseksi. Lihasten toiminnan edellytyksenä on supistuvien proteiinien lisäksi myös kalsiumionien läsnäolo soluissa. Siksi tätä alkuainetta sisältävien elintarvikkeiden riittämätön tai liiallinen kulutus voi johtaa lihasten virheelliseen toimintaan - sekä sileään että poikkijuovaiseen.
Lisäksi soluissa on toinen spesifinen proteiini - myoglobiini. On välttämätöntä sitoutua hapella ja varastoida se.
Mitä tulee organelleihin, lihaskudokselle on myofibrillien lisäksi erityistä solun suuren määrän mitokondrioita - soluhengityksestä vastaavia kaksoiskalvoorganelleja. Ja tämä ei ole yllättävää, koska lihassäikeet tarvitsevat suuren määrän energiaa supistuakseen, mitä mitokondriot tuottavat hengityksen aikana.

Joillakin myosyyteillä on myös useampi kuin yksi ydin. Tämä on tyypillistä poikkijuovaisille lihaksille, joiden solut voivat sisältää noin kaksikymmentä ydintä, ja joskus tämä luku saavuttaa sata. Tämä johtuu siitä, että poikkijuovainen lihaskuitu muodostuu useista soluista, jotka yhdistetään myöhemmin yhdeksi.
Poikkijuovaisten lihasten rakenne
Tämän tyyppistä kudosta kutsutaan myös luurankolihakseksi. Tämäntyyppisten lihasten kuidut ovat pitkiä, kerätty nippuihin. Niiden solujen pituus voi olla useita senttimetrejä (jopa 10-12). Ne sisältävät monia ytimiä, mitokondrioita ja myofibrillejä. Jokaisen poikkijuovaisen kudoksen myofibrillin perusrakenneyksikkö on sarkomeeri. Se koostuu supistuvista proteiineista.
Tämän lihaksen pääominaisuus on, että sitä voidaan hallita tietoisesti, toisin kuin sileät ja sydänlihakset.
Tämän kudoksen kuidut kiinnitetään luihin jänteiden avulla. Siksi tällaisia lihaksia kutsutaan luurankoiksi.
Sileän lihaskudoksen rakenne
Sileät lihakset reunustavat joitain sisäelimiä, kuten suolistoa, kohtua, virtsarakkoa ja verisuonia. Lisäksi niistä muodostuu sulkijalihaksia ja nivelsiteitä.

Sileä lihassyy ei ole yhtä pitkä kuin poikkijuovainen lihaskuitu. Mutta sen paksuus on suurempi kuin luurankolihasten tapauksessa. Sileät lihassolut ovat muodoltaan karan muotoisia pikemminkin kuin lankamainen muoto, kuten poikkijuovaiset lihassolut.
Sileiden lihasten supistumista välittäviä rakenteita kutsutaan protofibrilleiksi. Toisin kuin myofibrillit, niillä on yksinkertaisempi rakenne. Mutta materiaali, josta ne on rakennettu, on samat supistuvat proteiinit aktiini ja myosiini.
Sileiden lihasten myosyyteissä on myös vähemmän mitokondrioita kuin poikkijuovaisissa ja sydänsoluissa. Lisäksi ne sisältävät vain yhden ytimen.
Sydänlihaksen ominaisuudet
Jotkut tutkijat määrittelevät sen poikkijuovaisen lihaskudoksen alatyypiksi. Niiden kuidut ovat todellakin samanlaisia monella tapaa. Sydänsolut - kardiomyosyytit - sisältävät myös useita ytimiä, myofibrillejä ja suuren määrän mitokondrioita. Tämä kudos pystyy myös supistumaan paljon nopeammin ja vahvemmin kuin sileä lihas.
Kuitenkin tärkein piirre, joka erottaa sydänlihaksen poikkijuovaisesta lihaksesta, on se, että sitä ei voida hallita tietoisesti. Sen supistuminen tapahtuu vain automaattisesti, kuten sileiden lihasten tapauksessa.

Tyypillisten solujen lisäksi sydänkudoksessa on myös erittäviä kardiomyosyyttejä. Ne eivät sisällä myofibrillejä eivätkä supistu. Nämä solut ovat vastuussa atriopeptiinihormonin tuottamisesta, mikä on tarpeen verenpaineen säätelyyn ja veren tilavuuden säätelyyn.
Poikkijuovaisten lihasten toiminnot
Niiden päätehtävänä on liikuttaa kehoa avaruudessa. Se on myös kehon osien liikettä suhteessa toisiinsa.
Muita poikkijuovaisten lihasten toimintoja ovat asennon ylläpitäminen sekä veden ja suolojen varastointi. Lisäksi niillä on suojaava rooli, mikä koskee erityisesti vatsalihaksia, jotka estävät sisäelinten mekaanisia vaurioita.

Poikkijuovaisten lihasten toimintoihin voi kuulua myös lämpötilan säätely, sillä aktiivisen lihasten supistumisen aikana vapautuu huomattava määrä lämpöä. Tästä syystä lihakset alkavat täristä tahattomasti jäätyessään.
Sileän lihaskudoksen toiminnot
Tämäntyyppinen lihas suorittaa evakuointitoiminnon. Se johtuu siitä, että suolen sileät lihakset työntävät ulosteita paikkaan, jossa ne erittyvät kehosta. Tämä rooli ilmenee myös synnytyksen aikana, kun kohdun sileät lihakset työntävät sikiön ulos elimestä.

Sileän lihaskudoksen toiminnot eivät rajoitu tähän. Niiden sulkijalihas on myös tärkeä. Tämän tyyppisestä kudoksesta muodostuu erityisiä pyöreitä lihaksia, jotka voivat sulkeutua ja avautua. Sulkijalihakset ovat virtsateissä, suolistossa, mahalaukun ja ruokatorven välissä, sappirakossa ja pupillissa.
Toinen sileiden lihasten tärkeä rooli on nivelsidelaitteen muodostuminen. On tarpeen säilyttää sisäelinten oikea asento. Kun näiden lihasten sävy laskee, joissakin elimissä voi esiintyä prolapsia.
Tähän päättyvät sileän lihaskudoksen toiminnot.
Sydänlihaksen tarkoitus
Tässä ei periaatteessa ole mitään erityistä puhuttavaa. Tämän kudoksen tärkein ja ainoa tehtävä on varmistaa verenkierto kehossa.
Johtopäätös: erot kolmen lihaskudostyypin välillä
Tämän ongelman selventämiseksi esitämme taulukon:
| Sileä lihas | Poikkijuovaiset lihakset | Sydämen lihaskudos |
| Kutistuu automaattisesti | Voidaan hallita tietoisesti | Kutistuu automaattisesti |
| Solut ovat pitkänomaisia, karan muotoisia | Solut ovat pitkiä, rihmamaisia | Pitkänomaiset solut |
| Kuituja ei ole niputettu | Kuidut yhdistetään nipuiksi | Kuidut yhdistetään nipuiksi |
| Yksi ydin per solu | Useita ytimiä solussa | Useita ytimiä solussa |
| Suhteellisen pieni määrä mitokondrioita | Suuri määrä mitokondrioita | |
| Ei myofibrillejä | Myofibrillejä esiintyy | Siellä on myofibrillejä |
| Solut pystyvät jakautumaan | Kuidut eivät voi jakautua | Solut eivät voi jakautua |
| Supistuu hitaasti, heikosti, rytmisesti | Sopimukset nopeasti ja vahvasti | Supistuu nopeasti, voimakkaasti, rytmisesti |
| Line sisäelimet (suoli, kohtu, virtsarakko), muodostavat sulkijalihakset | Kiinnitetty luurankoon | Muotoile sydän |
Siinä kaikki poikkijuovaisen, sileän ja sydänlihaskudoksen tärkeimmät ominaisuudet. Nyt tunnet niiden toiminnot, rakenteen sekä tärkeimmät erot ja yhtäläisyydet.
Kuten luurankolihaksissa, laukaisin stimuloi useimpien sileiden lihasten supistumista on solunsisäisten kalsiumionien määrän kasvu. Erilaisissa sileissä lihaksissa tämä lisääntyminen voi johtua hermostimulaatiosta, hormonistimulaatiosta, kuidun venymisestä tai jopa kuitua ympäröivän ympäristön kemiallisen koostumuksen muutoksesta.
Kuitenkin sisään sileistä lihaksista puuttuu troponiini(säätelyproteiini, jonka kalsium aktivoi). Sileän lihaksen supistuminen aktivoituu täysin erilaisella mekanismilla, joka on kuvattu alla.
Kalsiumionien yhdistelmä kalmoduliinin kanssa. Myosiinikinaasin aktivointi ja myosiinipään fosforylaatio.
Troponiinin sijaan sileät lihassolut sisältävät suuria määriä toista säätelyproteiinia, kalmoduliinia. Vaikka tämä proteiini on samanlainen kuin troponiini, se eroaa tavasta, jolla se laukaisee supistumisen. Kalmoduliini tekee tämän aktivoimalla myosiinin ristisiltoja. Aktivointi ja vähentäminen suoritetaan seuraavassa järjestyksessä.
1. Kalsiumionit sitoutuvat kalmoduliiniin.
2. Kalmoduliini-kalsiumkompleksi sitoutuu fosforyloivaan entsyymiin myosiinikinaasiin ja aktivoi sen.
3. Yksi kunkin myosiinipään kevytketjuista, jota kutsutaan säätelyketjuksi, fosforyloituu myosiinikinaasin toimesta. Kun tämä ketju ei ole fosforyloitu, myosiinipään syklistä kiinnittymistä ja irtoamista aktiinifilamentin suhteen ei tapahdu. Mutta kun säätelyketju fosforyloituu, pää saa kyvyn sitoutua uudelleen aktiinifilamenttiin ja suorittaa koko syklisen jaksollisen "vedonlyönnin", joka on supistumisen taustalla, kuten luurankolihaksissa.
Vähennyksen lopettaminen. Myosiinifosfataasin rooli. Kun kalsiumionien pitoisuus putoaa kriittisen tason alapuolelle, edellä mainitut prosessit kehittyvät automaattisesti päinvastaiseen suuntaan myosiinipään fosforylaatiota lukuun ottamatta. Tämän tilan kehityksen kääntämiseksi tarvitaan toinen entsyymi - myosiinifosfataasia, joka sijaitsee sileän lihassolun nesteisiin ja katkaisee fosfataasin säätelykevytketjusta. Tämän jälkeen syklinen aktiivisuus ja siten supistuminen pysähtyy.
Siksi aika välttämätön lihasten rentoutumiseen, määräytyy suurelta osin aktiivisen myosiinifosfataasin määrästä solussa.
Mahdollinen mekanismi "salpa"-mekanismin säätämiseksi. Salpamekanismin tärkeydestä johtuen sileän lihaksen toiminnassa tätä ilmiötä yritetään selittää, koska sen avulla voidaan ylläpitää pitkäkestoista sileää lihasjännitystä monissa elimissä ilman merkittävää energiankulutusta. Monien ehdotettujen mekanismien joukossa esittelemme yhden yksinkertaisimmista.
Kun se aktivoituu voimakkaasti ja myosiinikinaasi, ja myosiinifosfataasi, myosiinipäiden kiertotaajuus ja supistumisnopeus ovat korkeat. Sitten, kun entsyymiaktivaatio laskee, syklin taajuus pienenee, mutta samaan aikaan näiden entsyymien deaktivoituminen sallii myosiinipäiden pysymisen kiinni aktiinifilamenteissa yhä pidempiä jaksoja. Näin ollen aktiinifilamenttiin kulloinkin kiinnitettyjen päiden lukumäärä pysyy suurena.
Numerosta lähtien päät kiinnitettynä aktiiniin määrittää supistumisen staattisen voiman, jännitystä pidetään yllä tai "salpataan". Energiaa kuluu kuitenkin vähän, koska ATP ei hajoa ADP:ksi, paitsi harvoissa tapauksissa, kun pää irrotetaan.
Sileä lihas niitä esiintyy ruoansulatuskanavan seinämissä, keuhkoputkissa, veressä ja imusuonissa, virtsarakossa, kohdussa sekä iiriksessä, sädelihaksessa, ihossa ja rauhasissa. Toisin kuin poikkijuovaiset lihakset, ne eivät ole erillisiä lihaksia, vaan muodostavat vain osan elimistä. Sileälihassoluilla on pitkänomainen kara- tai nauhamainen muoto, joissa on terävät päät. Niiden pituus ihmisillä on yleensä noin 20 mikronia. Sileät lihassolut saavuttavat suurimman pituuden (jopa 500 mikronia) raskaana olevan ihmisen kohdun seinämässä. Solun keskiosassa on sauvamainen ydin, ja sytoplasmassa koko solua pitkin kulkevat ohuet, täysin homogeeniset myofibrillet rinnakkain. Siksi solussa ei ole poikittaisia juovia. Paksummat myofibrillit sijaitsevat solun ulkokerroksissa. Niitä kutsutaan rajoilla ja niillä on yksiakselinen kahtaistaite. Elektronimikroskooppi osoittaa, että myofibrillit ovat protofibrillinippuja ja niissä on poikkijuovia, jotka eivät näy valomikroskoopissa. Sileät lihassolut voivat uusiutua jakautumalla (mitoosi). Ne sisältävät eräänlaista aktomyosiinia - tonoaktomyosiinia. Sileiden lihassolujen välillä on samat kalvokontaktialueet eli yhteydet kuin sydämen solujen välillä, joita pitkin virityksen ja eston oletetaan leviävän sileästä lihassolusta toiseen.
Sileissä lihaksissa viritys leviää hitaasti, sileiden lihasten supistukset johtuvat luustolihaksia vahvemmista ja pidempään kestävistä ärsykkeistä. Sen supistumisen piilevä jakso kestää useita sekunteja. Sileät lihakset supistuvat paljon hitaammin kuin luurankolihakset. Siten sammakon mahalaukun sileän lihaksen supistumisaika on 15-20 s. Sileät lihassupistukset voivat kestää useita minuutteja tai jopa tunteja. Toisin kuin luustolihakset, sileän lihaksen supistukset ovat tonisoivia. Sileät lihakset pystyvät olemaan pitkään tonisoivassa jännityksessä erittäin vähäisellä aine- ja energiankulutuksella. Esimerkiksi ruoansulatuskanavan, virtsarakon, sappirakon, kohdun ja muiden elinten sulkijalihasten sileät lihakset ovat hyvässä kunnossa kymmeniä minuutteja ja useita tunteja. Korkeampien selkärankaisten verisuonten seinämien sileät lihakset pysyvät hyvässä kunnossa läpi elämän.
Lihaksessa syntyvien impulssien taajuuden ja sen jännitystason välillä on suora yhteys. Mitä korkeampi taajuus, sitä suurempi on sävy tiettyyn rajaan asti, mikä johtuu ei-samanaikaisesti jännittyneiden lihaskuitujen jännitysten summasta.
Sileillä lihaksilla on maukkuus - kyky säilyttää pituus venytettynä muuttamatta jännitystä, toisin kuin luurankolihakset, jotka ovat jännittyneitä venytettäessä.
Toisin kuin luustolihakset, monet sileät lihakset ovat automaattisia. Ne supistuvat paikallisten refleksimekanismien, kuten ruoansulatuskanavan Meissner- ja Auerbach-punnuksien, tai verenkiertoon pääsevien kemikaalien, kuten asetyylikoliinin, norepinefriinin ja adrenaliinin, vaikutuksesta. Sileiden lihasten automaattiset supistukset tehostuvat tai estyvät hermojärjestelmästä tulevien hermoimpulssien vaikutuksesta. Siksi, toisin kuin luustolihaksissa, on olemassa erityisiä estäviä hermoja, jotka pysäyttävät supistumisen ja aiheuttavat sileän lihaksen rentoutumisen. Joillakin sileillä lihaksilla, joissa on suuri määrä hermopäätteitä, ei ole automaattisuutta, esimerkiksi pupillin sulkijalihaksessa, kissan nikottelevassa kalvossa.
Sileät lihakset voivat lyhentyä huomattavasti, paljon enemmän kuin luurankolihakset. Yksittäinen stimulaatio voi aiheuttaa sileän lihaksen supistumisen 45 % ja maksimaalinen supistuminen toistuvalla stimulaatiorytmillä voi olla 60-75 %.
Sileä lihaskudos kehittyy myös mesodermista (syntyy mesenkyymistä); se koostuu yksittäisistä, erittäin pitkänomaisista karan muotoisista soluista, jotka ovat kooltaan paljon pienempiä poikkijuovaisten lihasten säikeisiin verrattuna. Niiden pituus vaihtelee välillä 20-500 μ ja leveys 4-7 μ. Yleensä näissä soluissa on yksi pitkänomainen ydin, joka sijaitsee solun keskellä. Solun protoplasmassa kulkee pitkittäissuunnassa lukuisia ja erittäin ohuita myofibrillejä, joissa ei ole poikittaisia juovia ja jotka ovat täysin näkymättömiä ilman erityiskäsittelyä. Jokainen sileä lihassolu on peitetty ohuella sidekudoskalvolla. Nämä kalvot yhdistävät naapurisolut toisiinsa. Toisin kuin poikkijuovaiset kuidut, jotka sijaitsevat lähes koko luurankolihaksen pituudelta, kaikissa sileälihaskompleksissa on huomattava määrä soluja, jotka sijaitsevat yhdessä linjassa.
Sileät lihassolut löytyvät kehosta joko hajallaan sidekudoksessa tai liittyneenä erikokoisiksi lihaskomplekseiksi.
Jälkimmäisessä tapauksessa jokaista lihassolua ympäröi joka puolelta myös solujen välinen aine, jonka läpi kulkevat hienoimmat fibrillit, joiden lukumäärä voi olla hyvinkin erilainen. Hienoimmat elastisten kuitujen verkostot löytyvät myös solujen välisestä aineesta.
Elinten sileät lihassolut yhdistyvät lihaskimpuiksi. Monissa tapauksissa (virtsatiet, kohtu jne.) nämä kimput haarautuvat ja sulautuvat muihin nippuihin muodostaen tiheydeltään vaihtelevia pintaverkkoja. Jos suuri määrä nippuja sijaitsee lähellä, muodostuu tiheä lihaskerros (esimerkiksi maha-suolikanava). Verensyöttö sileille lihaksille suoritetaan suonten kautta, jotka kulkevat suurten sidekudoskerrosten läpi nippujen välillä; kapillaarit tunkeutuvat kunkin nipun kuitujen väliin ja muodostavat sitä pitkin haarautuneena tiheän kapillaariverkoston. Sileä lihaskudos sisältää myös imusuonet. Sileitä lihaksia hermottavat autonomisen hermoston kuidut. Sileät lihassolut, toisin kuin poikkijuovaiset lihaskuidut, tuottavat hitaita, jatkuvia supistuksia. He pystyvät työskentelemään pitkään ja suurella voimalla. Esimerkiksi kohdun lihaksikkaat seinämät kehittävät tuntikausia kestävän synnytyksen aikana voimaa, johon poikkijuovaiset lihakset eivät pääse käsiksi. Sileiden lihasten toiminta ei pääsääntöisesti ole tahtomme alainen (kasvihermotus, katso alla) - ne ovat tahattomia.
Sileälihas on kehitykseltään (fylogenia) muinaisempaa kuin poikkijuovalihas, ja se on yleisempi eläinmaailman alemmissa muodoissa.
Sileiden lihasten luokitus
Sileät lihakset jaetaan viskeraalisiin (yksittäisiin) ja monimuotoisiin lihaksiin. Viskeraalisia sileitä lihaksia löytyy kaikista sisäelimistä, ruuansulatusrauhasten kanavista, verestä ja imusuonista sekä ihosta. Monimuotoisia lihaksia ovat sädelihas ja iirislihas. Sileiden lihasten jako viskeraalisiin ja monimuotoisiin lihaksiin perustuu niiden motorisen hermotuksen erilaisiin tiheyksiin. Viskeraalisessa sileässä lihaksessa motorisia hermopäätteitä on pienessä määrässä sileälihassoluja. Tästä huolimatta hermopäätteiden viritys välittyy kaikkiin nipun sileisiin lihassoluihin johtuen viereisten myosyyttien - yhteyksien - välisistä tiiviistä kosketuksista. Nexes sallii toimintapotentiaalien ja hitaita depolarisaation aaltoja levitä lihassolusta toiseen, joten sisäelinten sileät lihakset supistuvat samanaikaisesti hermoimpulssin saapuessa.
Sileiden lihasten toiminnot ja ominaisuudet
Muovi. Toinen tärkeä sileän lihaksen erityisominaisuus on jännityksen vaihtelevuus ilman säännöllistä yhteyttä sen pituuteen. Siten jos sisäelinten sileää lihasta venytetään, sen jännitys kasvaa, mutta jos lihas pidetään venytyksen aiheuttamassa venymistilassa, jännitys vähenee vähitellen, toisinaan ei vain tasolle, joka oli ennen venytystä, vaan myös tämän tason alapuolella. Tätä ominaisuutta kutsutaan sileän lihaksen plastisuudesta. Siten sileä lihas muistuttaa enemmän viskoosia muovimassaa kuin huonosti taipuisaa rakenteellista kudosta. Sileiden lihasten plastisuus edistää onttojen sisäelinten normaalia toimintaa.
Kiihtymisen ja supistumisen suhde. Sähköisten ja mekaanisten ilmenemismuotojen välistä suhdetta on vaikeampi tutkia sisäelinten sileässä lihaksessa kuin luuranko- tai sydänlihaksessa, koska sisäelinten sileä lihas on jatkuvassa toiminnan tilassa. Suhteellisen levon olosuhteissa voidaan tallentaa yksi AP. Sekä luuston että sileän lihaksen supistuminen perustuu aktiinin liukumiseen suhteessa myosiiniin, jossa Ca2+-ioni suorittaa laukaisevan toiminnon.
Sileiden lihasten supistumismekanismilla on ominaisuus, joka erottaa sen luurankolihasten supistumismekanismista. Tämä ominaisuus on, että ennen kuin sileän lihaksen myosiini voi osoittaa ATPaasiaktiivisuuttaan, se on fosforyloitava. Myosiinin fosforylaatiota ja defosforylaatiota havaitaan myös luurankolihaksissa, mutta siinä fosforylaatioprosessi ei ole välttämätön myosiinin ATPaasiaktiivisuuden aktivoimiseksi. Sileiden lihasten myosiinin fosforylaatiomekanismi on seuraava: Ca2+-ioni yhdistyy kalmoduliiniin (kalmoduliini on Ca2+-ionin vastaanottava proteiini). Tuloksena oleva kompleksi aktivoi entsyymin, myosiinin kevytketjukinaasin, joka puolestaan katalysoi myosiinin fosforylaatioprosessia. Sitten aktiini liukuu myosiinia vastaan, joka muodostaa supistumisen perustan. Huomaa, että sileän lihaksen supistumisen laukaisee Ca2+-ionin lisääminen kalmoduliiniin, kun taas luusto- ja sydänlihaksissa laukaisee Ca2+:n lisääminen troponiiniin.
Kemiallinen herkkyys. Sileät lihakset ovat erittäin herkkiä erilaisille fysiologisesti aktiivisille aineille: adrenaliinille, norepinefriinille, ACh:lle, histamiinille jne. Tämä johtuu spesifisten reseptoreiden läsnäolosta sileän lihaksen solukalvolla. Jos suolen sileän lihaksen valmisteeseen lisätään adrenaliinia tai norepinefriiniä, kalvopotentiaali kasvaa, AP:n taajuus vähenee ja lihas rentoutuu, eli havaitaan sama vaikutus kuin sympaattisten hermojen kiihtyessä.
Norepinefriini vaikuttaa sileän lihaksen solukalvon α- ja β-adrenergisiin reseptoreihin. Norepinefriinin vuorovaikutus β-reseptoreiden kanssa alentaa lihasten sävyä adenylaattisyklaasin aktivoitumisen ja syklisen AMP:n muodostumisen ja sitä seuraavan solunsisäisen Ca2+:n sitoutumisen lisääntymisen seurauksena. Norepinefriinin vaikutus α-reseptoreihin estää supistumista lisäämällä Ca2+-ionien vapautumista lihassoluista.
ACh vaikuttaa kalvopotentiaaliin ja suoliston sileän lihaksen supistumiseen, mikä on päinvastainen kuin norepinefriinin vaikutus. ACh:n lisääminen suolen sileän lihaksen valmisteeseen vähentää kalvopotentiaalia ja lisää spontaanien AP:iden esiintymistiheyttä. Tämän seurauksena sävy kohoaa ja rytmiset supistukset lisääntyvät, eli havaitaan sama vaikutus kuin parasympaattisten hermojen kiihtyessä. ACh depolarisoi kalvon ja lisää sen Na+:n ja Ca+:n läpäisevyyttä.
Joidenkin elinten sileät lihakset reagoivat erilaisiin hormoneihin. Siten kohdun sileät lihakset eläimillä ovulaation ja munasarjojen poistamisen välisenä aikana ovat suhteellisen kiihtymättömiä. Kiiman aikana tai eläimillä, joille on poistettu munasarjat, joille on annettu estrogeenia, sileän lihaksen kiihtyvyys lisääntyy. Progesteroni lisää kalvopotentiaalia jopa enemmän kuin estrogeeni, mutta tässä tapauksessa kohdun lihasten sähköinen ja supistuva toiminta estyy.
Sileät lihakset ovat osa sisäelimiä. Supistumisen ansiosta ne tarjoavat elinten (ruoansulatuskanavan, virtsaelimen, verisuonten jne.) motorisen toiminnan. Toisin kuin luustolihakset, sileät lihakset ovat tahattomia.
Morfo-funktionaalinen rakenne sileä lihaksia. Sileän lihaksen päärakenneyksikkö on lihassolu, joka on karan muotoinen ja on ulkopuolelta peitetty plasmakalvolla. Elektronimikroskoopin alla kalvossa voidaan nähdä lukuisia syvennyksiä - caveolae, jotka lisäävät merkittävästi lihassolun kokonaispintaa. Lihassolun sarkolemmat sisältää plasmakalvon sekä sitä ulkopuolelta peittävän tyvikalvon ja viereiset kollageenisäikeet. Tärkeimmät solunsisäiset elementit: ydin, mitokondriot, lysosomit, mikrotubulukset, sarkoplasminen verkkokalvo ja supistuvat proteiinit.
Lihassolut muodostavat lihaskimppuja ja lihaskerroksia. Solujen välinen tila (100 nm tai enemmän) on täynnä elastisia ja kollageenikuituja, kapillaareja, fibroblasteja jne. Joillakin alueilla viereisten solujen kalvot ovat erittäin tiukasti (solujen välinen rako 2-3 nm). Oletetaan, että nämä alueet (nexus) palvelevat solujen välistä viestintää ja virityksen välittämistä. On todistettu, että joissakin sileissä lihaksissa on suuri määrä sidoksia (pupillisulkijalihakset, ohutsuolen pyöreät lihakset jne.), kun taas toisissa niitä on vähän tai ei ollenkaan (vas deferens, suolen pituussuuntaiset lihakset). Nahtamattomien lihassolujen välillä on myös väli- tai desmopodibny-yhteys (kalvon paksuuntumisen ja soluprosessien avulla). Ilmeisesti nämä yhteydet ovat tärkeitä solujen mekaaniselle kytkennälle ja mekaanisen voiman siirtämiselle solujen kautta.
Myosiinin ja aktiinin protofibrillien kaoottisen jakautumisen vuoksi sileät lihassolut eivät ole poikkijuovaisia, kuten luusto- ja sydänsolut. Toisin kuin luustolihaksissa, sileissä lihaksissa ei ole T-järjestelmää, ja sarkoplasminen retikulumi muodostaa vain 2-7 % myoplasman tilavuudesta, eikä sillä ole yhteyksiä solun ulkoiseen ympäristöön.
Sileiden lihasten fysiologiset ominaisuudet .
Sileät lihassolut, kuten poikkijuovaiset, supistuvat aktiiniprotofibrillien liukumisen vuoksi myosiiniprotofibrillien välillä, mutta ATP:n liukumis- ja hydrolyysinopeus ja siten supistumisnopeus on 100-1000 kertaa pienempi kuin poikkijuovaisissa lihaksissa. Tämän ansiosta sileät lihakset ovat hyvin sopeutuneet pitkäaikaiseen luistoon vähäisellä energiankulutuksella ja ilman väsymystä.
Sileät lihakset, ottaen huomioon kyvyn tuottaa AP:tä vasteena kynnys- tai supra-sarvi-stimulaatioon, jaetaan perinteisesti faasillisiin ja tonic-stimulaatioihin. Faasilihakset synnyttävät täysimittaisen potentiaalisen toiminnan, kun taas tonic lihakset tuottavat vain paikallisen, vaikka niillä on myös mekanismi täysimittaisten potentiaalien luomiseksi. Tonic-lihasten kyvyttömyys suorittaa AP:ta selittyy kalvon korkealla kaliumin läpäisevyydellä, mikä estää regeneratiivisen depolarisaation kehittymisen.
Nahtamattomien lihasten sileiden lihassolujen kalvopotentiaalin arvo vaihtelee välillä -50 - -60 mV. Kuten muissakin lihaksissa, hermosolut mukaan lukien, pääasiassa +, Na +, Cl- osallistuvat sen muodostumiseen. Ruoansulatuskanavan, kohdun ja joidenkin verisuonten sileissä lihassoluissa kalvopotentiaali on epävakaa, spontaaneja heilahteluja havaitaan hitaiden depolarisaatioaaltojen muodossa, joiden huipulla voi ilmaantua AP-purkauksia. Sileiden lihasten toimintapotentiaalin kesto vaihtelee välillä 20-25 ms - 1 s tai enemmän (esimerkiksi virtsarakon lihaksissa), ts. se on pidempi kuin luurankolihasten AP:n kesto. Sileiden lihasten toimintamekanismissa Na +:n vieressä Ca2 +:lla on tärkeä rooli.
Spontaani myogeeninen aktiivisuus. Toisin kuin luustolihaksilla, mahalaukun, suoliston, kohdun ja virtsanjohtimien sileillä lihaksilla on spontaania myogeenistä toimintaa, ts. kehittää spontaaneja tetanohyodiinisupistuksia. Ne varastoidaan olosuhteissa, joissa nämä lihakset on eristetty ja intrafusaaliset hermoplexukset on farmakologisesti kytketty pois päältä. Joten AP esiintyy itse sileissä lihaksissa, eikä sitä aiheuta hermoimpulssien siirtyminen lihaksiin.
Tämä spontaani aktiivisuus on myogeenistä alkuperää ja sitä esiintyy lihassoluissa, jotka toimivat sydämentahdistimena. Näissä soluissa paikallinen potentiaali saavuttaa kriittisen tason ja siirtyy AP:hen. Mutta kalvon repolarisaation jälkeen syntyy spontaanisti uusi paikallinen potentiaali, joka aiheuttaa toisen AP:n jne. AP, joka leviää nexuksen kautta viereisiin lihassoluihin nopeudella 0,05-0,1 m/s, peittää koko lihaksen aiheuttaen sen supistumisen. Esimerkiksi vatsan peristalttisia supistuksia esiintyy taajuudella 3 kertaa minuutissa, segmentaalisia ja heilurimaisia paksusuolen liikkeitä - 20 kertaa minuutissa yläosissa ja 5-10 kertaa minuutissa alemmissa osissa. Siten näiden sisäelinten sileillä lihaskuiduilla on automaattisuutta, mikä ilmenee niiden kyvystä supistua rytmisesti ulkoisten ärsykkeiden puuttuessa.
Mikä on syy potentiaalin ilmaantumiseen tahdistimen sileissä lihassoluissa? Ilmeisesti se johtuu kaliumin vähenemisestä ja kalvon natriumin ja kalsiumin läpäisevyyden lisääntymisestä. Mitä tulee säännölliseen esiintymiseen hitaiden depolarisaation aaltojen osalta, jotka ovat selkeimpiä maha-suolikanavan lihaksissa, niiden ionisesta alkuperästä ei ole luotettavaa tietoa. Ehkä tietty rooli on kaliumvirran alkuperäisen inaktivoivan komponentin vähenemisellä lihassolujen depolarisaation aikana vastaavien kalium-ionikanavien inaktivoitumisen vuoksi.
Sileiden lihasten elastisuus ja venyvyys. Toisin kuin luustolihakset, sileät lihakset toimivat venytettyinä plastisina, joustavina rakenteina. Muovisuuden ansiosta sileät lihakset voidaan rentoutua täysin sekä supistuneessa että venytetyssä tilassa. Esimerkiksi mahalaukun tai virtsarakon seinämän sileiden lihasten plastisuus näiden elinten täyttyessä estää onkalonsisäisen paineen nousun. Liiallinen venyttely johtaa usein supistuksen stimulaatioon, joka johtuu sydämentahdistinsolujen depolarisaatiosta, joka tapahtuu lihasta venytettäessä, ja siihen liittyy toimintapotentiaalin lisääntyminen ja sen seurauksena supistumisen lisääntyminen. Venytysprosessia aktivoivalla supistumisella on suuri rooli verisuonten tyviäänen itsesäätelyssä.
Sileiden lihasten supistumisen mekanismi. Edellytyksenä esiintymiselle on sileiden lihasten sekä luustolihasten supistuminen ja Ca2 + -pitoisuuden nousu myoplasmassa (jopa 10-5 M). Uskotaan, että supistumisprosessia aktivoi ensisijaisesti solunulkoinen Ca2+, joka tulee lihassoluihin jänniteohjattujen Ca2+-kanavien kautta.
Sileiden lihasten neuromuskulaarisen transmission erityispiirre on, että hermotuksen suorittaa autonominen hermosto ja sillä voi olla sekä kiihottavaa että estävää vaikutusta. Tyypin mukaan on kolinergisiä (välittäjä asetyylikoliini) ja adrenergisiä (välittäjänorepinefriini) välittäjiä. Ensimmäiset löytyvät yleensä ruoansulatusjärjestelmän lihaksista, jälkimmäiset verisuonten lihaksista.
Sama lähetin joissakin synapseissa voi olla kiihottava ja toisissa inhiboiva (riippuen sytoreseptoreiden ominaisuuksista). Adrenergiset reseptorit jaetaan a- ja b-. α-adrenergisiin reseptoreihin vaikuttava norepinefriini supistaa verisuonia ja estää ruuansulatuskanavan motiliteettia ja B-adrenergisiin reseptoreihin vaikuttava stimuloi sydämen toimintaa ja laajentaa joidenkin elinten verisuonia, rentouttaa keuhkoputkien lihaksia. . Kuvattu neuromuskulaarinen-. siirtyminen sileissä lihaksissa muiden välittäjien avulla.
Vasteena eksitatorisen lähettimen vaikutukselle tapahtuu sileiden lihassolujen depolarisaatiota, joka ilmenee eksitatorisen synaptisen potentiaalin (ESP) muodossa. Kun se saavuttaa kriittisen tason, PD tapahtuu. Tämä tapahtuu, kun useita impulsseja lähestyy hermopäätettä peräkkäin. SMM:n esiintyminen on seurausta postsynaptisen kalvon Na +:n, Ca2+:n ja SI:n läpäisevyyden lisääntymisestä."
Inhiboiva lähetin aiheuttaa postsynaptisen kalvon hyperpolarisaation, joka ilmenee estävässä synaptisessa potentiaalissa (ISP). Hyperpolarisaatio perustuu kalvon läpäisevyyden lisääntymiseen, pääasiassa K+:n osalta. Inhiboivan välittäjän rooli asetyylikoliinin kiihottamissa sileissä lihaksissa (esim. suolen lihakset, keuhkoputket) on norepinefriinilla, ja sileissä lihaksissa, joissa norepinefriini on kiihottava välittäjä (esimerkiksi virtsarakon lihakset), asetyylikoliini pelaa. rooli.
Kliininen ja fysiologinen puoli. Joissakin sairauksissa, kun luustolihasten hermotus häiriintyy, niiden passiiviseen venytykseen tai siirtymiseen liittyy niiden sävyn refleksin nousu, ts. vastustuskyky venymiselle (spastisuus tai jäykkyys).
Jos verenkierto on heikentynyt, sekä tiettyjen aineenvaihduntatuotteiden (maito- ja fosforihappo), myrkyllisten aineiden, alkoholin, väsymyksen tai lihasten lämpötilan laskun alaisena (esimerkiksi pitkittyneen kylmän veden uinnin aikana), kontraktuura voi esiintyä esiintyy pitkäaikaisen aktiivisen lihassupistuksen jälkeen. Mitä enemmän lihasten toiminta on heikentynyt, sitä selvempi on kontraktuurin jälkivaikutus (esim. puremislihasten supistumisleuan alueen patologiassa). Mikä on kontraktuurin alkuperä? Uskotaan, että kontraktuuri syntyi ATP:n pitoisuuden laskusta lihaksessa, mikä johti pysyvän yhteyden muodostumiseen ristisiltojen ja aktiiniprotofibrillien välille. Tässä tapauksessa lihas menettää joustavuutta ja kovettuu. Kontraktuuri poistuu ja lihas rentoutuu, kun ATP-pitoisuus saavuttaa normaalin tason.
Sairaudissa, kuten myotoniassa, lihassolukalvot kiihtyvät niin helposti, että pienikin ärsytys (esimerkiksi neulaelektrodin asettaminen elektromyografian aikana) aiheuttaa lihasimpulssien purkauksen. Spontaanit AP:t (värinäpotentiaalit) kirjataan myös ensimmäisessä vaiheessa lihaksen denervaation jälkeen (kunnes toimettomuus johtaa sen surkastumiseen).
Lihaskudos
Tehokas hermotus Sileää lihaskudosta hoitavat autonomisen hermoston sekä sympaattinen (noradrenerginen hermotus) että parasympaattinen (kolinerginen hermotus) osa, joilla on päinvastainen vaikutus lihaskudoksen supistumisaktiivisuuteen. Sen serotonerginen ja peptiderginen hermotus on myös kuvattu. Hermopäätteitä löytyy vain yksittäisistä soluista, ja ne näyttävät suonikohjuilta ohuilta aksonihaaruksilta. Viritys välittyy viereisiin myosyytteihin rakoliitosten kautta.
Afferentti hermotus sen tarjoavat hermosäikeiden oksat, jotka muodostavat vapaita päitä sileään lihaskudokseen.
Sileän lihaskudoksen toiminnan humoraalinen säätely. Hormonit ja muut biologisesti aktiiviset aineet vaikuttavat sileän lihaskudoksen (erilaisittain eri elimissä) supistumisaktiivisuuteen, koska sen soluissa on vastaavia reseptorisarjoja. Näitä aineita ovat histamiini, serotoniini, bradykiniini, endoteliini, typpioksidi, leukotrieenit, prostaglandiinit, neurotensiini, substanssi P, kolekystokiniini, vasoaktiini interstiinipeptidi (VIP), opioidit jne. Kohdun myosyyttien supistukset raskauden lopussa ja synnytyksen aikana stimuloidaan hormonin oksitosiini toimesta; estrogeeni lisääntyy ja progesteroni vähentää niiden sävyä.
Sileän lihaskudoksen myogeeninen aktiivisuus. Sileiden myoniittien fysiologinen ärsyke on niiden venyminen, mikä aiheuttaa sarkolemman depolarisaation ja Ca 2+ -ionien sisäänvirtauksen sarkoplasmaan. Sileälle lihaskudokselle on ominaista spontaani rytminen aktiivisuus (automaattisuus), joka johtuu sarkolemman kalsiumpumppujen syklisesti muuttuvasta aktiivisuudesta. Spontaani aktiivisuus on voimakkainta suolen, kohdun ja virtsateiden sileässä lihaskudoksessa; se on paljon heikompaa verisuonten lihaskudoksessa. Automaatiolle tyypillisimmät syklit ovat supistuminen ja rentoutuminen, joiden kesto on keskimäärin noin 1 minuutti. (0,5 - 2 minuuttia). Normaaleissa olosuhteissa tähän myogeeniseen toiminnan rytmiin vaikuttavat hermo- ja hormonaaliset signaalit, jotka vahvistavat, heikentävät, koordinoivat ja synkronoivat myosyyttien supistumisaktiivisuutta.
Sileän lihaskudoksen fysiologinen regeneraatio suoritetaan jatkuvasti subsellulaarisella tasolla päivittämällä solukomponentteja.
Sileän lihaksen hypertrofia toimii sen reaktiona toiminnallisen kuormituksen lisääntymiseen, joka yleensä liittyy sen venymiseen.
HERMOKUDOSTO
Hermokudos koostuu hermosoluista (neurosyytit, hermosolut itse), joilla on kyky tuottaa ja johtaa hermoimpulsseja, ja neurogliasoluista, jotka suorittavat useita aputoimintoja (tuki-, trofia-, este-, suoja- jne.) ja varmistavat neuronien toimintaa. Neuronit ja neuroglia (lukuun ottamatta yhtä sen lajikkeista, mikrogliaa) ovat hermosolujen johdannaisia.
NEURONIT
Neuronit (neurosyytit, itse hermosolut) ovat erikokoisia soluja (jotka vaihtelevat kehon pienimmistä - hermosoluista, joiden kehon halkaisija on 4-5 mikronia - suurimpiin, joiden kehon halkaisija on noin 140 mikronia). Niiden kokonaismäärä ihmisen hermostossa on yli 100 miljardia (10 11) ja joidenkin arvioiden mukaan biljoona (10 12). Syntyessään hermosolut menettävät kykynsä jakautua, joten syntymän jälkeisen elämän aikana niiden lukumäärä ei kasva, vaan päinvastoin, solujen luonnollisen menetyksen vuoksi vähenee vähitellen.
Sileälihas on supistuva kudos, joka koostuu yksittäisistä soluista ja ilman poikittaisia juovia (kuva 1.). Sileä lihassolu on muodoltaan karan muotoinen, noin 50-400 µm pitkä ja 2-10 µm paksu. Yksittäiset langat yhdistetään erityisillä solujen välisillä kontakteilla - desmosomeilla, ja ne muodostavat verkoston, johon on kudottu kollageenikuituja. Sydän- ja luustolihaksille tyypillisten poikkijuovien puuttuminen selittyy myosiini- ja aktiinifilamenttien epäsäännöllisellä jakautumisella. Sileät lihakset lyhenevät myös myofilamenttien liukumisen vuoksi toisiinsa nähden, mutta liukumisnopeus ja ATP:n hajoaminen on täällä 100 - 1000 kertaa pienempi kuin poikkijuovaisten lihasten. Sileät lihakset ovat tässä suhteessa erityisen hyvin sopeutuneet pitkäaikaiseen kestävään supistukseen, mikä ei johda väsymykseen ja merkittävään energiankulutukseen.
Sileät lihakset ovat osa sisäelimiä, verisuonia ja ihoa. Ne erottuvat mielenkiintoisista toiminnallisista ominaisuuksista: kyvystä suorittaa suhteellisen hitaita liikkeitä ja pitkittyneitä tonic-supistuksia. Hitaat liikkeet (supistukset), joilla on usein onttojen elinten seinämien sileiden lihasten rytminen supistuminen: vatsa, suolet, ruoansulatusrauhasten kanavat, virtsarakko, sappirakko, varmistavat näiden elinten sisällön liikkeen. Esimerkkinä ovat suoliston heiluri- ja peristalttiset liikkeet. Sileiden lihasten pitkittyneet tonisoidut supistukset ovat erityisen voimakkaita onttojen elinten sulkijalihaksissa; niiden tonic supistukset estävät sisällön vapautumisen. Tämä varmistaa sapen esiintymisen sappirakossa ja virtsan esiintymisen virtsarakossa sekä ulosteiden muodostumisen paksusuolessa.
Näyttää selkärankaisten poikkijuovaisten ja sileiden lihasten rakenteen (vasemmalla) sekä sähköisen (yhtenäiset viivat) ja mekaanisen (katkoviivat) toiminnan (oikealla) välisen suhteen. A. Poikkijuovaiset lihakset ovat monitumaisia sylinterimäisiä soluja. Ne synnyttävät nopeita toimintapotentiaalia ja nopeita supistuksia. B. Sileillä lihaskuiduilla on yksi ydin, pieni koko ja fusiform muoto. Ne on liitetty toisiinsa sivupinnoillaan rakoliitosten kautta ja muodostavat sähköisesti yhtenäisiä soluryhmiä.
Hermotus on diffuusi, kuitujen aktivointi tapahtuu välittäjän vapautumisen vuoksi autonomista hermoa pitkin sijaitsevista jatkeista. Vaikka sileiden lihassolujen toimintapotentiaalit ovat nopeita, tuloksena olevat supistukset ovat hitaita ja pitkäkestoisia.
Verisuonten seinämien ohuet sileät lihakset, erityisesti valtimot ja valtimot, ovat jatkuvassa tonisoivassa supistuksessa. Valtimoiden seinämien lihaskerroksen sävy säätelee verenpainetta ja verenkiertoa elimiin.
Sileiden lihasten motorisen hermotuksen suorittavat autonomisen hermoston solujen prosessit, herkät - sympaattisten ganglioiden solujen prosessit. Sileiden lihasten sävyä ja motorista toimintaa säätelevät myös humoraaliset vaikutukset.
Kaikki sileät lihakset voidaan jakaa kahteen ryhmään:
1. Sileät lihakset myogeenisellä aktiivisuudella. Monissa suoliston sileissä lihaksissa (esim. umpisuolessa) yksittäinen toimintapotentiaalin aiheuttama supistuminen kestää useita sekunteja. Tämän seurauksena supistukset, jotka seuraavat alle 2 s:n välein, menevät päällekkäin, ja yli 1 Hz:n taajuudella sulautuvat enemmän tai vähemmän tasaiseksi tetanukseksi (jäykkäkouristusmainen sävy) (kuva 2). Tällaisen tetanuksen luonne on myogeeninen; Toisin kuin luustolihakset, suolen, virtsanjohtimen, mahalaukun ja kohdun sileät lihakset kykenevät spontaaneihin teetaanin kaltaisiin supistuksiin eristämisen ja denervaation jälkeen ja jopa intramuraalisten ganglionihermosolujen salpauksen jälkeen. Näin ollen niiden toimintapotentiaalit eivät johdu hermoimpulssien siirtymisestä lihakseen, vaan ne ovat myogeenistä alkuperää.
Myogeeninen viritys tapahtuu sydämentahdistinsoluissa, jotka ovat rakenteeltaan identtisiä muiden lihassolujen kanssa, mutta eroavat sähköfysiologisista ominaisuuksista. Tahdistimen potentiaalit depolarisoivat kalvon kynnystasolle aiheuttaen toimintapotentiaalin. Kationien (pääasiassa Ca2+) sisäänpääsyn vuoksi kalvo depolarisoituu nollatasolle ja jopa muuttaa polariteetin +20 mV:iin muutaman millisekunnin ajaksi. Repolarisaation jälkeen seuraa uusi sydämentahdistinpotentiaali, joka varmistaa seuraavan toimintapotentiaalin muodostumisen. Kun paksusuolen valmiste altistetaan asetyylikoliinille, sydämentahdistinsolut depolarisoituvat lähelle kynnystasoa ja toimintapotentiaalien tiheys kasvaa. Niiden aiheuttamat supistukset sulautuvat lähes sileäksi tetanukseksi. Mitä korkeampi toimintapotentiaalien taajuus, sitä yhtenäisempi jäykkäkouristus ja sitä voimakkaampi on yksittäisten supistusten summauksesta johtuva supistuminen. Sitä vastoin noradrenaliinin levittäminen samaan valmisteeseen muodostaa hyperpolaarisen kalvon ja sen seurauksena vähentää toimintapotentiaalien esiintymistiheyttä ja tetanuksen suuruutta. Nämä ovat mekanismeja, joilla autonominen hermosto ja sen välittäjät moduloivat tahdistimen spontaanin toiminnan.

Kuva 2.
Asetyylikoliinihoito (nuoli) lisää toimintapotentiaalien taajuutta niin, että yksittäiset lyönnit sulautuvat tetanukseksi. Alin ennätys on lihasjännityksen aika.
2. Sileät lihakset ilman myogeenistä toimintaa. Toisin kuin suoliston lihakset, valtimoiden, siementiehyiden, iiriksen ja sädelihasten sileillä lihaksilla on yleensä vähän tai ei ollenkaan spontaania toimintaa. Niiden supistuminen tapahtuu näihin lihaksiin autonomisten hermojen kautta toimitettujen impulssien vaikutuksesta. Tällaiset ominaisuudet johtuvat niiden kudoksen rakenteellisesta organisoinnista. Vaikka siinä olevat solut ovat sähköisesti yhteydessä toisiinsa, monet niistä muodostavat suoria synaptisia kontakteja niitä hermottavien aksonien kanssa, mutta eivät muodosta tavanomaisia hermo-lihassynapseja sileässä lihaskudoksessa. Lähettimen vapautuminen tapahtuu lukuisista paksunnuksista (jatkeista), jotka sijaitsevat autonomisten aksonien pituudella (kuva 1).
Välittäjät saavuttavat lihassolut diffuusion kautta ja aktivoivat ne. Samaan aikaan soluissa syntyy virityspotentiaalia, jotka muuttuvat toimintapotentiaaliksi, jotka aiheuttavat tetaanisen supistumisen.
Sileiden lihasten toiminnot ja ominaisuudet
Sähköinen toiminta. Viskeraalisille sileille lihaksille on ominaista epävakaa kalvopotentiaali. Kalvopotentiaalin vaihtelut hermostovaikutuksista riippumatta aiheuttavat epäsäännöllisiä supistuksia, jotka pitävät lihaksen jatkuvassa osittaisen supistumisen tilassa - tonuksessa. Sileiden lihasten sävy näkyy selvästi onttojen elinten sulkijalihaksissa: sappirakko, rakko, mahalaukun ja pohjukaissuolen ja ohutsuolen paksusuolen liitoskohdassa sekä pienten valtimoiden ja valtimoiden sileissä lihaksissa. arteriolit. Sileiden lihassolujen kalvopotentiaali ei heijasta lepopotentiaalin todellista arvoa. Kun kalvopotentiaali pienenee, lihas supistuu, ja kun kalvopotentiaali kasvaa, se rentoutuu.
Automaatio. Sileiden lihassolujen toimintapotentiaalit ovat luonteeltaan autorhytmisiä (tahdistin), jotka ovat samanlaisia kuin sydämen johtumisjärjestelmän potentiaalit. Sydämentahdistimen potentiaalit rekisteröidään sileän lihaksen eri alueilla. Tämä osoittaa, että kaikki sisäelinten sileät lihassolut kykenevät spontaanisti automaattiseen toimintaan. Sileiden lihasten automaattisuus, ts. kyky automaattiseen (spontaaniin) toimintaan on luontainen monille sisäelimille ja verisuonille.
Vetovaste. Viskeraalisen sileän lihaksen ainutlaatuinen ominaisuus on sen reaktio venytykseen. Vasteena venytykseen sileät lihakset supistuvat. Tämä johtuu siitä, että venyttely vähentää solujen kalvopotentiaalia, lisää AP:n frekvenssiä ja viime kädessä sileiden lihasten sävyä. Ihmiskehossa tämä sileän lihaksen ominaisuus toimii yhtenä keinona säädellä sisäelinten motorista toimintaa. Esimerkiksi, kun vatsa on täynnä, sen seinämä venyy. Vatsan seinämän sävyn lisääntyminen sen venymisen seurauksena auttaa ylläpitämään elimen tilavuutta ja sen seinämien paremman kosketuksen saapuvan ruoan kanssa. Verisuonissa verenpaineen vaihteluiden aiheuttama turvotus on tärkeä tekijä verisuonten sävyn myogeenisessä itsesäätelyssä. Lopuksi, kasvavan sikiön aiheuttama kohdun lihasten venyttely on yksi syistä synnytyksen alkamiseen.
Muovi. Toinen tärkeä sileän lihaksen erityisominaisuus on jännityksen vaihtelevuus ilman säännöllistä yhteyttä sen pituuteen. Siten jos sisäelinten sileää lihasta venytetään, sen jännitys kasvaa, mutta jos lihas pidetään venytyksen aiheuttamassa venymistilassa, jännitys vähenee vähitellen, toisinaan ei vain tasolle, joka oli ennen venytystä, vaan myös tämän tason alapuolella. Tätä ominaisuutta kutsutaan sileän lihaksen plastisuudesta. Siten sileä lihas muistuttaa enemmän viskoosia muovimassaa kuin huonosti taipuisaa rakenteellista kudosta. Sileiden lihasten plastisuus edistää onttojen sisäelinten normaalia toimintaa.
Kiihtymisen ja supistumisen suhde. Sähköisten ja mekaanisten ilmenemismuotojen välistä suhdetta on vaikeampi tutkia sisäelinten sileässä lihaksessa kuin luuranko- tai sydänlihaksessa, koska sisäelinten sileä lihas on jatkuvassa toiminnan tilassa. Suhteellisen levon olosuhteissa voidaan tallentaa yksi AP. Sekä luuston että sileän lihaksen supistuminen perustuu aktiinin liukumiseen suhteessa myosiiniin, jossa Ca2+-ioni suorittaa laukaisevan toiminnon.
Sileiden lihasten supistumismekanismilla on ominaisuus, joka erottaa sen luurankolihasten supistumismekanismista. Tämä ominaisuus on, että ennen kuin sileän lihaksen myosiini voi osoittaa ATPaasiaktiivisuuttaan, se on fosforyloitava. Myosiinin fosforylaatiota ja defosforylaatiota havaitaan myös luurankolihaksissa, mutta siinä fosforylaatioprosessi ei ole välttämätön myosiinin ATPaasiaktiivisuuden aktivoimiseksi.
Kemiallinen herkkyys. Sileät lihakset ovat erittäin herkkiä erilaisille fysiologisesti aktiivisille aineille: adrenaliinille, norepinefriinille, ACh:lle, histamiinille jne. Tämä johtuu spesifisten reseptoreiden läsnäolosta sileän lihaksen solukalvolla. Jos suolen sileän lihaksen valmisteeseen lisätään adrenaliinia tai norepinefriiniä, kalvopotentiaali kasvaa, AP:n taajuus vähenee ja lihas rentoutuu, eli havaitaan sama vaikutus kuin sympaattisten hermojen kiihtyessä.
Norepinefriini vaikuttaa sileiden lihassolujen kalvon b- ja b-adrenergisiin reseptoreihin. Norepinefriinin vuorovaikutus β-reseptoreiden kanssa alentaa lihasten sävyä adenylaattisyklaasin aktivoitumisen ja syklisen AMP:n muodostumisen ja sitä seuraavan solunsisäisen Ca2+:n sitoutumisen lisääntymisen seurauksena. Norepinefriinin vaikutus β-reseptoreihin estää supistumista lisäämällä Ca2+-ionien vapautumista lihassoluista.
ACh vaikuttaa kalvopotentiaaliin ja suoliston sileän lihaksen supistumiseen, mikä on päinvastainen kuin norepinefriinin vaikutus. ACh:n lisääminen suolen sileän lihaksen valmisteeseen vähentää kalvopotentiaalia ja lisää spontaanien AP:iden esiintymistiheyttä. Tämän seurauksena sävy kohoaa ja rytmiset supistukset lisääntyvät, eli havaitaan sama vaikutus kuin parasympaattisten hermojen kiihtyessä. ACh depolarisoi kalvon ja lisää sen Na+:n ja Ca+:n läpäisevyyttä.
Joidenkin elinten sileät lihakset reagoivat erilaisiin hormoneihin. Siten kohdun sileät lihakset eläimillä ovulaation ja munasarjojen poistamisen välisenä aikana ovat suhteellisen kiihtymättömiä. Kiiman aikana tai eläimillä, joille on poistettu munasarjat, joille on annettu estrogeenia, sileän lihaksen kiihtyvyys lisääntyy. Progesteroni lisää kalvopotentiaalia jopa enemmän kuin estrogeeni, mutta tässä tapauksessa kohdun lihasten sähköinen ja supistuva toiminta estyy.