این فرستنده اصلی نورون های خودمختار در نظر گرفته می شود. سیستم عصبی خودمختار (ANS)
به آنها هسته می گویند. آنها به عنوان حلقه های اتصال بین ساختارهای سیستم عصبی عمل می کنند، پردازش اولیه تکانه ها را انجام می دهند و مسئول عملکرد اندام های احشایی هستند.
بدن انسان دو نوع عملکرد را انجام می دهد - و رویشی. سوماتیک شامل درک محرک های خارجی و پاسخ به آنها با استفاده از ماهیچه های اسکلتی است. این واکنش ها توسط هوشیاری انسان قابل کنترل است و سیستم عصبی مرکزی مسئولیت اجرای آنها را بر عهده دارد.
عملکردهای رویشی - هضم، متابولیسم، خون سازی، گردش خون، تنفس، تعریق و غیره - توسط بدن کنترل می شود که به آگاهی انسان بستگی ندارد. سیستم خودمختار علاوه بر تنظیم کار اندام های احشایی، تروفیسم را برای عضلات و سیستم عصبی مرکزی فراهم می کند.
عقده های مسئول عملکردهای جسمی نشان دهنده گره های نخاعی و گره های عصبی جمجمه ای هستند. اتونومیک بسته به محل مراکز در سیستم عصبی مرکزی به دو دسته پاراسمپاتیک و سمپاتیک تقسیم می شود.
اولی ها در دیواره های اندام قرار دارند و سمپاتیک ها از راه دور در ساختاری به نام تنه مرزی قرار دارند.
ساختار گانگلیون
بسته به ویژگی های مورفولوژیکی، اندازه گانگلیون ها از چند میکرومتر تا چند سانتی متر متغیر است. در اصل، مجموعه ای از سلول های عصبی و گلیال است که با یک غشای همبند پوشانده شده است.
عنصر بافت همبند توسط عروق لنفاوی و خونی نفوذ می کند. هر سلول عصبی (یا گروهی از نوروسیت ها) توسط یک غشای کپسولی احاطه شده است که در داخل با اندوتلیوم و از خارج با رشته های بافت همبند پوشیده شده است. در داخل کپسول یک سلول عصبی و ساختارهای گلیال وجود دارد که عملکرد نورون را تضمین می کند.
یک آکسون منفرد که با یک غلاف میلین پوشیده شده است از نورون خارج می شود که به دو قسمت منشعب می شود. یکی از آنها بخشی از عصب محیطی است و یک گیرنده را تشکیل می دهد و دومی به سیستم عصبی مرکزی فرستاده می شود.
مراکز خودمختار در ساقه مغز و نخاع قرار دارند. مراکز پاراسمپاتیک در ناحیه جمجمه و ساکرال و مراکز سمپاتیک در ناحیه توراکولومبار قرار دارند.
گانگلیون سیستم عصبی خودمختار
 سیستم سمپاتیک شامل دو نوع گره است: مهره ای و پیش مهره ای.
سیستم سمپاتیک شامل دو نوع گره است: مهره ای و پیش مهره ای.
مهره ها در دو طرف ستون فقرات قرار دارند و تنه های مرزی را تشکیل می دهند. آنها توسط رشته های عصبی به نخاع متصل می شوند که شاخه های سفید و خاکستری اتصال را ایجاد می کنند. رشته های عصبی خارج شده از گره به سمت اندام های احشایی هدایت می شوند.
پیش مهره ایدر فاصله بیشتری از ستون فقرات قرار دارند، در حالی که آنها همچنین در فاصله ای از اندام هایی که مسئولیت آنها را بر عهده دارند قرار دارند. نمونه هایی از گره های پیش مهره ای عبارتند از: خوشه های گردنی، مزانتریک نورون ها و شبکه خورشیدی.
پاراسمپاتیکاین بخش توسط عقده های واقع در اندام ها یا در مجاورت آنها تشکیل می شود.
شبکه عصبی داخل ارگانیروی اندام یا دیواره آن قرار دارد. شبکه های داخل اندامی بزرگ در عضله قلب، در لایه عضلانی دیواره روده و در پارانشیم اندام های غده قرار دارند.
گانگلیون های سیستم عصبی خودمختار و مرکزی دارای خواص زیر هستند:
- هدایت سیگنال در یک جهت؛
- الیافی که وارد گره می شوند با مناطق تأثیر یکدیگر همپوشانی دارند.
- جمع فضایی (مجموع تکانه ها می تواند یک پتانسیل در سلول عصبی ایجاد کند).
- انسداد (تحریک اعصاب باعث پاسخ کمتری نسبت به تحریک هر عصب جداگانه می شود).
تأخیر سیناپسی در گانگلیون های اتونوم بیشتر از ساختارهای مشابه سیستم عصبی مرکزی است و پتانسیل پس سیناپسی طولانی است. موجی از تحریک در سلول های عصبی گانگلیونی با افسردگی جایگزین می شود. این عوامل در مقایسه با سیستم عصبی مرکزی منجر به ریتم ضربه نسبتاً پایینی میشوند.
گانگلیون ها چه وظایفی را انجام می دهند؟
هدف اصلی گره های اتونوم توزیع و انتقال تکانه های عصبی و همچنین تولید رفلکس های موضعی است. هر گانگلیون، بسته به موقعیت و ویژگی های تغذیه ای خود، مسئول عملکرد یک ناحیه خاص از بدن است.
 گانگلیون ها با استقلال از سیستم عصبی مرکزی مشخص می شوند که به آنها اجازه می دهد تا فعالیت اندام ها را بدون مشارکت مغز و نخاع تنظیم کنند.
گانگلیون ها با استقلال از سیستم عصبی مرکزی مشخص می شوند که به آنها اجازه می دهد تا فعالیت اندام ها را بدون مشارکت مغز و نخاع تنظیم کنند.
ساختار گره های داخل دیواره شامل سلول های ضربان ساز است که می تواند فرکانس انقباضات ماهیچه های صاف روده را تنظیم کند.
این ویژگی با قطع فیبرهای CNS است که به اندام های داخلی در گره های محیطی سیستم خودمختار هدایت می شوند، جایی که سیناپس ها را تشکیل می دهند. در این مورد، آکسون های خارج شده از گانگلیون به طور مستقیم بر اندام داخلی تأثیر می گذارند.
هر فیبر عصبی که وارد گانگلیون سمپاتیک می شود تا سی سلول عصبی پس گانگلیونی را عصب دهی می کند. این باعث می شود سیگنال را چند برابر کرده و تکانه تحریک را از گانگلیون عصبی پخش کند.
در گره های پاراسمپاتیک، یک فیبر بیش از چهار نوروسیت را عصب دهی می کند و انتقال تکانه به صورت موضعی اتفاق می افتد.
گانگلیون - مراکز رفلکس
عقده های سیستم عصبی در قوس رفلکس شرکت می کنند که امکان تصحیح فعالیت اندام ها و بافت ها را بدون مشارکت مغز فراهم می کند. در پایان قرن نوزدهم، بافت شناس روسی، دوگل، در نتیجه آزمایش هایی که روی شبکه های عصبی در دستگاه گوارش مطالعه می کرد، سه نوع نورون - حرکتی، بینابینی و گیرنده و همچنین سیناپس های بین آنها را شناسایی کرد.
 وجود سلولهای عصبی گیرنده نیز امکان پیوند عضله قلب را از اهداکننده به گیرنده تأیید میکند. اگر تنظیم ضربان قلب از طریق سیستم عصبی مرکزی انجام شود، پس از پیوند قلب، سلولهای عصبی دچار انحطاط میشوند. نورون ها و سیناپس ها در اندام پیوندی به کار خود ادامه می دهند که نشان دهنده استقلال آنهاست.
وجود سلولهای عصبی گیرنده نیز امکان پیوند عضله قلب را از اهداکننده به گیرنده تأیید میکند. اگر تنظیم ضربان قلب از طریق سیستم عصبی مرکزی انجام شود، پس از پیوند قلب، سلولهای عصبی دچار انحطاط میشوند. نورون ها و سیناپس ها در اندام پیوندی به کار خود ادامه می دهند که نشان دهنده استقلال آنهاست.
در پایان قرن بیستم، مکانیسمهای رفلکسهای محیطی که گرههای رویشی پیش مهرهای و درون دیوارهای را میسازند، به طور تجربی ایجاد شدند. توانایی ایجاد قوس بازتابی مشخصه برخی گره ها است.
رفلکس های محلی به شما امکان می دهد سیستم عصبی مرکزی را تسکین دهید، تنظیم عملکردهای مهم را قابل اعتمادتر کنید و در صورت قطع ارتباط با سیستم عصبی مرکزی قادر به ادامه عملکرد مستقل اندام های داخلی باشید.
گره های خودمختار اطلاعات مربوط به عملکرد اندام ها را دریافت و پردازش می کنند و سپس آن را به مغز می فرستند. این باعث ایجاد یک قوس بازتابی در هر دو سیستم خودمختار و جسمی می شود که نه تنها رفلکس ها، بلکه واکنش های رفتاری آگاهانه را نیز تحریک می کند.
جزئیات
گانگلیوننمایندگی کند خوشه های نورون های چند قطبی (یک آکسون و چند دندریت).(از چند سلول تا ده ها هزار). عقده های خارج ارگانی (سمپاتیک) دارای یک کپسول بافت همبند کاملاً مشخص به عنوان ادامه پری نوریوم هستند. عقده های پاراسمپاتیک معمولاً در شبکه های عصبی داخل دیواره قرار دارند. عقدههای شبکههای داخل دیواره، مانند سایر عقدههای خودمختار، حاوی نورونهای خودمختار کمانهای رفلکس موضعی هستند. نورون های چند قطبی با قطر 20-35 میکرومتر به صورت پراکنده قرار دارند، هر نورون توسط گلیوسیت های گانگلیونی احاطه شده است.
علاوه بر آن شرح داده شده است نورونهای غدد درونریز، گیرندههای شیمیایی، دوقطبی و در برخی از مهرهداران، نورونهای تک قطبی. گانگلیون های سمپاتیک حاوی سلول های کوچک و به شدت فلورسنت (سلول های MYF) با فرآیندهای کوتاه و تعداد زیادی وزیکول دانه ای در سیتوپلاسم هستند. آنها کاتکول آمین ها را آزاد می کنند و اثر مهاری بر انتقال تکانه ها از رشته های عصبی پیش گانگلیونی به نورون سمپاتیک وابران دارند. به این سلول ها، اینترنورون می گویند.
در میان رشته های اصلی نورون های چند قطبیرویشی گانگلیونمتمایز: موتور (سلول های دوگل نوع 1)، حساس (سلول های دوگل نوع II) و انجمنی (سلول های دوگل نوع III). نورون های حرکتی دارای دندریت های کوتاه با امتداد لایه ای ("پدهای گیرنده") هستند. آکسون این سلول ها بسیار طولانی است، به عنوان بخشی از رشته های عصبی نازک بدون میلین پس از گانگلیون فراتر از گانگلیون می رود و به میوسیت های صاف اندام های داخلی ختم می شود. سلول های نوع 1 نورون های آکسون بلند نامیده می شوند. نورون های نوع دوم سلول های عصبی متساوی الاضلاع هستند. 2-4 فرآیند از بدن آنها گسترش می یابد که در میان آنها تشخیص آکسون دشوار است. بدون انشعاب، فرآیندها به دور از بدن نورون گسترش می یابند. دندریت های آنها دارای انتهای عصبی حسی است و آکسون به بدنه نورون های حرکتی در گانگلیون های مجاور ختم می شود. سلولهای نوع II نورونهای حساس قوسهای بازتابی اتونوم محلی هستند. سلولهای Dogel نوع III از نظر شکل بدن شبیه به نورونهای خودمختار نوع II هستند، اما دندریتهای آنها فراتر از گانگلیون گسترش نمییابند و نوریت به سمت سایر گانگلیونها هدایت میشود. بسیاری از محققان این سلول ها را نوعی نورون حسی می دانند.
بنابراین، در گانگلیون های اتونوم محیطی، کمان های رفلکس محلی متشکل از نورون های اتونوم حسی، حرکتی و احتمالاً انجمنی وجود دارد.
گانگلیون های اتونوم داخل دیواره در دیواره دستگاه گوارش با این واقعیت متمایز می شوند که در ترکیب آنها، علاوه بر نورون های کولینرژیک حرکتی، نورون های مهاری نیز وجود دارد. آنها توسط سلول های عصبی آدرنرژیک و پورینرژیک نشان داده می شوند. در دومی، واسطه یک نوکلئوتید پورین است. در گانگلیون های اتونوم داخل دیواره نیز نورون های پپتیدرژیک وجود دارد که پپتید عروقی روده ای، سوماتوستاتین و تعدادی از پپتیدهای دیگر را ترشح می کنند که با کمک آنها تنظیم نورواندوکرین و تعدیل فعالیت بافت ها و اندام های دستگاه گوارش انجام می شود.
استیل کولین- گیرنده های نیکوتین (بلوک کورار، هگزامتونیوم)، موسکارینی (بلوک آتروپین). فعال سازی گیرنده → تولید EPSP. EPSP سریع (N-cholinocerus)→ باز کردن کانال های یونی. EPSP آهسته (M-cholinorets) ← سرکوب جریان M ناشی از افزایش رسانایی K.
نوروپپتیدها- به عنوان تعدیل کننده عصبی عمل کنید.
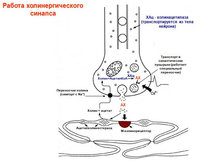
انکفالین ها، ماده P، لولیبرین، نوروتنسین، سوماتوستاتین – علامت. گانگلیون (+Ach)
کاتکولامین ها(NA، دوپامین) انتقال دهنده های عصبی سلول های کوچک با فلورسانس شدید هستند.
نوروپپتید Y، سوماتوستاتین - علامت. پست گانگلیونی
پستگانگلیونی های سمپاتیک: NA، ATP، نوروپپتید U.
α1→ اینوزوتول تری فسفات، دی اسیل گلیسرول. فعال سازی پروتئین α2→G، ↓cAMP.
β → پروتئین G → AC → cAMP

استثناها: واسطه درد، گیرنده های موسکارینی.
پاراسمپ postganglionics: Ach، VIP، NO، سوماتوستاتین، ATP، پپتیدهای مخدر.
M1 (میل ترکیبی بالا برای پیرنزپین) - ترشح اسید توسط سلول های غدد معده را افزایش می دهد، M2 (کم) - ضربان قلب را کاهش می دهد. ریتم، ترشح غدد اشکی و بزاقی.
اقدام متنوع:
- ثانیه خاص واسطه ها: M2 می تواند بر روی IP3 عمل کند یا می تواند روی AC عمل کند و cAMP را کاهش دهد.
-عمل در کانال های K و Ca
- NO روی اندوتلیوم → گوانیلات سیکلاز → cGMP → پروتئین کیناز وابسته به cGMP → شل شدن عضلات صاف تشکیل می شود.
در سیستم عصبی خودمختارتشخیص بخش مرکزی و محیطی بخش های مرکزی سیستم عصبی سمپاتیک توسط هسته های شاخ های جانبی طناب نخاعی سینه ای نشان داده می شود. در سیستم عصبی پاراسمپاتیک، تقسیمات مرکزی شامل هستههای مغز میانی و بصل النخاع و همچنین هستههای شاخهای جانبی نخاع خاجی است. رشته های پاراسمپاتیک ناحیه کرانیوبولبار به عنوان بخشی از جفت های III، VII، IX و X اعصاب جمجمه ای ظاهر می شوند.
قسمت های محیطی سیستم عصبی خودمختارتوسط تنه های عصبی، گانگلیون ها و شبکه ها تشکیل می شود.
قوس های بازتابی اتونومیکبا یک نورون حسی شروع می شود که بدن آن مانند قوس های رفلکس جسمی در گانگلیون نخاعی قرار دارد. نورون های انجمن در شاخ های جانبی نخاع قرار دارند. در اینجا، تکانههای عصبی به نورونهای پیشگانگلیونی میانی تغییر میکنند که فرآیندهای آنها هستههای مرکزی را ترک میکنند و به عقدههای خودمختار میرسند، جایی که تکانهها را به نورون حرکتی منتقل میکنند. در این راستا، رشته های عصبی پیش گانگلیونی و پس گانگلیونی متمایز می شوند. اولین آنها سیستم عصبی مرکزی را به عنوان بخشی از ریشه های شکمی اعصاب نخاعی و اعصاب جمجمه ای ترک می کنند. در هر دو سیستم سمپاتیک و پاراسمپاتیک، رشته های عصبی پیشگانگلیونی متعلق به نورون های کولینرژیک هستند. آکسون های نورون های واقع در گانگلیون های خودمختار را پس گانگلیونی می نامند. آنها تماس مستقیم با سلول های موثر ایجاد نمی کنند. بخش های انتهایی آنها در طول مسیر انبساط هایی را تشکیل می دهند - واریکوزیته ها که حاوی حباب های واسطه هستند. در ناحیه وریدهای واریسی، غشای گلیال وجود ندارد و انتقال دهنده عصبی که در محیط منتشر می شود، بر سلول های موثر (به عنوان مثال، سلول های غدد، میوسیت های صاف و غیره) تأثیر می گذارد.
در گانگلیون های محیطیسیستم عصبی سمپاتیک، به عنوان یک قاعده، حاوی نورونهای وابران آدرنرژیک است (به استثنای نورونهایی که با غدد عرق ارتباط سیناپسی دارند، جایی که نورونهای سمپاتیک کولینرژیک هستند). در گانگلیون های پاراسمپاتیک، نورون های وابران همیشه کولینرژیک هستند.
گانگلیونخوشه هایی از نورون های چند قطبی (از چند سلول تا ده ها هزار). عقده های خارج ارگانی (سمپاتیک) دارای یک کپسول بافت همبند کاملاً مشخص به عنوان ادامه پری نوریوم هستند. عقده های پاراسمپاتیک معمولاً در شبکه های عصبی داخل دیواره قرار دارند. عقدههای شبکههای داخل دیواره، مانند سایر عقدههای خودمختار، حاوی نورونهای خودمختار کمانهای رفلکس موضعی هستند. نورون های چند قطبی با قطر 20-35 میکرومتر به صورت پراکنده قرار دارند، هر نورون توسط گلیوسیت های گانگلیونی احاطه شده است. علاوه بر این، نورونهای عصبی غدد درونریز، گیرندههای شیمیایی، دوقطبی و در برخی از مهرهداران، نورونهای تک قطبی توصیف شدهاند. گانگلیون های سمپاتیک حاوی سلول های کوچک و به شدت فلورسنت (سلول های MYF) با فرآیندهای کوتاه و تعداد زیادی وزیکول دانه ای در سیتوپلاسم هستند. آنها کاتکول آمین ها را آزاد می کنند و اثر مهاری بر انتقال تکانه ها از رشته های عصبی پیش گانگلیونی به نورون سمپاتیک وابران دارند. به این سلول ها، اینترنورون می گویند.
در میان نورون های چند قطبی بزرگعقده های اتونومیک متمایز می شوند: حرکتی (سلول های دوگل نوع I)، حساس (سلول های دوگل نوع دوم) و انجمنی (سلول های دوگل نوع III). نورون های حرکتی دارای دندریت های کوتاه با امتداد لایه ای ("پدهای گیرنده") هستند. آکسون این سلول ها بسیار طولانی است، به عنوان بخشی از رشته های عصبی نازک بدون میلین پس از گانگلیون فراتر از گانگلیون می رود و به میوسیت های صاف اندام های داخلی ختم می شود. سلول های نوع I را نورون های آکسون بلند می نامند. نورون های نوع دوم سلول های عصبی متساوی الاضلاع هستند. 2-4 فرآیند از بدن آنها گسترش می یابد که در میان آنها تشخیص آکسون دشوار است. بدون انشعاب، فرآیندها به دور از بدن نورون گسترش می یابند. دندریت های آنها دارای انتهای عصبی حسی است و آکسون به بدنه نورون های حرکتی در گانگلیون های مجاور ختم می شود. سلولهای نوع II نورونهای حساس قوسهای بازتابی اتونوم محلی هستند. سلولهای Dogel نوع III از نظر شکل بدن شبیه به نورونهای خودمختار نوع II هستند، اما دندریتهای آنها فراتر از گانگلیون گسترش نمییابند و نوریت به سمت سایر گانگلیونها هدایت میشود. بسیاری از محققان این سلول ها را نوعی نورون حسی می دانند.
بنابراین، در گانگلیون اتونوم محیطیقوسهای انعکاسی موضعی متشکل از نورونهای خودمختار حسی، حرکتی و احتمالاً انجمنی وجود دارد.
گانگلیون های اتونوم داخل دیوارهدر دیواره دستگاه گوارش متفاوت است که در ترکیب آنها، علاوه بر نورون های کولینرژیک حرکتی، نورون های مهاری نیز وجود دارد. آنها توسط سلول های عصبی آدرنرژیک و پورینرژیک نشان داده می شوند. در دومی، واسطه یک نوکلئوتید پورین است. در گانگلیون های اتونوم داخل دیواره نیز نورون های پپتیدرژیک وجود دارد که پپتید عروقی روده ای، سوماتوستاتین و تعدادی از پپتیدهای دیگر را ترشح می کنند که با کمک آنها تنظیم نورواندوکرین و تعدیل فعالیت بافت ها و اندام های دستگاه گوارش انجام می شود.
فیلم آموزشی آناتومی سیستم عصبی خودمختار (ANS)
اگر در تماشا کردن مشکل دارید، ویدیو را از صفحه دانلود کنید4. عقده های خودمختار، خواص آنها. مفهوم سیستم عصبی متاسمپاتیک و واسطه های آن.
یکی از ویژگی های بخش محیطی سیستم عصبی خودمختار وجود گانگلیون ها است که مجموعه ای از نورون ها هستند.
عقده های اتونومیک نقش مهمی در توزیع و انتشار تأثیرات عصبی بر روی اندام ها دارند. اشاره شده است که تعداد سلول های عصبی در گانگلیون چندین برابر تعداد رشته های پیش گانگلیونی است.
پدیده همگرایی در گانگلیون مشاهده می شود. در همان زمان، پدیده جمع مکانی و زمانی آشکار می شود. عقده های خودمختار همان ویژگی های مراکز عصبی سوماتیک را نشان می دهند. بنابراین، گانگلیون های سیستم عصبی خودمختار را گاهی مراکز عصبی می نامند که در حاشیه قرار دارند.
متاسمپاتیکسیستم عصبی (درون ارگانی) (MNS) مجموعه ای از تشکیلات عصبی است - نورون ها که بدن آنها عقده ها و فرآیندهای سلول های عصبی را فراتر از گانگلیون تشکیل می دهند. این ساختارها در دیواره قلب، روده و سایر اندام ها قرار دارند. تعداد نورون های این سیستم از تعداد نورون های نخاع بیشتر است. MNF بخش مرکزی ندارد، یعنی نسبتاً مستقل است. ماژول عملکردی آن شامل یک ضربان ساز، سلول های حسی، نورون های بینابینی، تونیک و عامل است. این تشکیلات عصبی استقلال اندام و تنظیم موضعی عملکرد ماهیچه های صاف و قلبی، اپیتلیوم ترشحی، دستگاه جذب و عروق خونی کوچک را فراهم می کنند. نقش سیستم عصبی متاسمپاتیک به ویژه در تنظیم عملکرد روده (بالای رکتوم)، جایی که تأثیرات عصبی مرکزی عملاً وجود ندارد، مهم است. حدود 20 واسطه و تعدیل کننده در سیناپس های MNS، از جمله استیل کولین، کوله سیستوکینین، انکفالین ها، هیستامین، سروتونین، سوماتوستاتین، ATP، ماده P، کاتکول آمین ها یافت شد. اعصاب سمپاتیک و پاراسمپاتیک می توانند سیناپس هایی را روی نورون های متاسمپاتیک ایجاد کنند و بر فعالیت آنها تأثیر بگذارند.
بلیط شماره 8
1. رفلکس به عنوان اصل اصلی سیستم عصبی مرکزی. مراحل اصلی مطالعه رفلکس. اختلاط معکوس، اهمیت آن برای بدن.
رفلکس(R.) یک واکنش طبیعی بدن به تغییرات در محیط خارجی یا داخلی است که با مشارکت سیستم عصبی در پاسخ به تحریک گیرنده ها رخ می دهد. کمان بازتاب- مسیر عصبی رفلکس - شامل یک پایانه عصبی حساس (یا سلول گیرنده)، یک رشته عصبی حسی با یک گانگلیون، یک بخش مرکزی (نرونهای حسی، بینابینی، عامل در سطوح مختلف سیستم عصبی مرکزی)، یک عصب وابران است. فیبر و یک عامل آموزه های رفلکسفیلسوف، ریاضیدان و فیزیولوژیست فرانسوی رنه دکارت (1648) به عنوان پاسخی به تحریکی که توسط مراکز عصبی نخاع تحقق می یابد ظاهر شد. او دو مفاد مهم نظریه رفلکس را تدوین کرد: 1) فعالیت بدن تحت تأثیر خارجی منعکس می شود (بعداً آن را رفلکس نامیدند: lat. reflexus - منعکس شده). 2) پاسخ به تحریک با استفاده از سیستم عصبی انجام می شود اصطلاح "رفلکس" برای اولین بار توسط فیزیولوژیست، آناتومیست و چشم پزشک چک I. Prochazka استفاده شد و عبارت "قوس رفلکس" توسط نوروپاتولوژیست انگلیسی و فیزیولوژیست M استفاده شد. سالن گام جدیدی در توسعه دکترین رفلکس کتاب "بازتاب های مغز" اثر I.M. Sechenov (1863) بود که ایده اصلی آن این بود: "همه اعمال زندگی خودآگاه و ناخودآگاه بازتاب هستند." به عبارت دیگر، I.M. Sechenov از اصل رفلکس برای توضیح مکانیسم های فعالیت مغز از جمله فرآیندهای تفکر استفاده کرد. این دانشمند عدم وجود پاسخ قابل مشاهده به عمل محرک ها را در تعدادی از موارد با ایجاد بازداری مرکزی توضیح داد که یک سال قبل (1862) کشف کرده بود. بنابراین، رفلکس ها ممکن است "پایان کوتاه" داشته باشند. پ. پاولوف که شاگرد مستقیم I. M. Sechenov نبود او را معلم خود می دانست و از اهمیت کتاب I. M. Sechenov بسیار قدردانی می کرد و آن را "یک ضربه درخشان از تفکر روسی" نامید.
I. P. Pavlov بیش از 30 سال از زندگی خود را به مطالعه رفلکس های بالاتر مغز اختصاص داد و برای این منظور از روش رفلکس های شرطی و بنابراین رویکردی عینی برای مطالعه عملکردهای مغز استفاده کرد. او یک نظریه بازتابی را بر اساس سه اصل ایجاد کرد: 1) جبرگرایی، یعنی علیت فرآیندهای مختلف فعالیت مغز. 2) تجزیه و تحلیل و سنتز محرک ها در قسمت های بالاتر مغز. 3) پیوند دینامیک با ساختار، به عنوان مثال، اتصال عملکردهای مغز با برخی از ساختارهای آن. مرحله فعلی توسعه دکترین رفلکس را می توان سیستمیک-سایبرنتیک نامید و با نام فیزیولوژیست های شوروی مرتبط است. A. Bernstein و P.K. Anokhin. ایده های بازخورد برای توضیح مکانیسم های فعالیت رفلکس توسط N. A. Bernstein (1947) استفاده شد که در نتیجه اصطلاح "حلقه رفلکس" ظاهر شد.
P.K. Anokhin (1949) تماس گرفت بازخورد رفلکس ها با "اختلاط معکوس". منبع آن گیرنده هایی است که در اندام مؤثر (1) و در اندام های حسی قرار دارند که در ارزیابی نتیجه یک عمل رفلکس شرکت می کنند (2). بنابراین، هنگام نواختن یک ملودی روی یک آلت موسیقی، چنین گیرنده هایی می توانند گیرنده های عمقی باشند ماهیچه ها و تاندون های دست (1)، و همچنین گیرنده های شنوایی (2). سیگنال ها اختلاس معکوسبرای مقایسه نتیجه یک عمل رفلکس با برنامه آن استفاده می شود.
در شرایط طبیعی زندگی، معمولاً رفلکس ها هستند
در سیستم ها متحد شده اند. علاوه بر این، عامل تشکیل دهنده سیستم نتیجه کلی است که اجرای این مجموعه از رفلکس ها به آن منجر می شود. بنابراین، حفظ غلظت مطلوب اکسیژن در پلاسمای خون توسط رفلکس های قلبی، تنفسی، حرکتی و سایر رفلکس ها که یک سیستم رفلکس عملکردی را تشکیل می دهند، تضمین می شود. دکترین سیستم های عملکردی برای تنظیم عملکردها توسط P.K. Anokhin (1949) ایجاد شد.
سیستم عصبی مرکزی انسان فعالیت های بدن را کنترل می کند و به چندین بخش تقسیم می شود. مغز سیگنال هایی را از بدن می فرستد و دریافت می کند و پس از پردازش آنها اطلاعاتی در مورد فرآیندها دارد. سیستم عصبی به دو سیستم عصبی خودمختار و سوماتیک تقسیم می شود.
تفاوت بین سیستم عصبی خودمختار و سوماتیک
سیستم عصبی سوماتیکتوسط هوشیاری انسان تنظیم می شود و می تواند فعالیت ماهیچه های اسکلتی را کنترل کند. تمام اجزای واکنش فرد به عوامل خارجی تحت کنترل نیمکره های مغز است. این واکنش های حسی و حرکتی انسان را فراهم می کند و تحریک و مهار آنها را کنترل می کند.
سامانه ی عصبی خودمختارفعالیت محیطی بدن را کنترل می کند و توسط هوشیاری کنترل نمی شود. در غیاب کامل هوشیاری با خودمختاری و اثرات عمومی بر بدن مشخص می شود. عصب دهی وابران اندام های داخلی به آن اجازه می دهد تا فرآیندهای متابولیک در بدن را کنترل کند و فرآیندهای تغذیه ای را برای عضلات اسکلتی، گیرنده ها، پوست و اندام های داخلی فراهم کند.
ساختار سیستم رویشی
سیستم عصبی خودمختار توسط هیپوتالاموس که در سیستم عصبی مرکزی قرار دارد کنترل می شود. سیستم عصبی خودمختار دارای ساختار متاسگمنتال است. مراکز آن در مغز، نخاع و قشر مغز قرار دارند. بخش های محیطی توسط تنه ها، گانگلیون ها و شبکه ها تشکیل می شوند.
سیستم عصبی خودمختار به دو دسته تقسیم می شود:
- دلسوز. مرکز آن در طناب نخاعی توراکولومبار قرار دارد. با گانگلیون های پاراورتبرال و پیش مهره ای ANS مشخص می شود.
- پاراسمپاتیک مراکز آن در مغز میانی و بصل النخاع، قسمت خاجی نخاع متمرکز شده است. بیشتر درون مدرسه ای
- متاسمپاتیک. دستگاه گوارش، رگ های خونی و اندام های داخلی بدن را عصب دهی می کند.

آن شامل:
- هسته های مراکز عصبی واقع در مغز و نخاع.
- عقده های اتونومیک که در امتداد محیط قرار دارند.
قوس بازتابی سیستم عصبی خودمختار
قوس بازتابی سیستم عصبی خودمختار از سه بخش تشکیل شده است:
- حساس یا آوران؛
- بینابینی یا انجمنی؛
- عامل

تعامل آنها بدون مشارکت نورون های اضافی مانند قوس بازتابی سیستم عصبی مرکزی اتفاق می افتد.
لینک حساس
واحد حسی در گانگلیون ستون فقرات قرار دارد. این گانگلیون دارای سلول های عصبی است که به صورت گروهی تشکیل شده اند و کنترل آنها توسط هسته های مرکزی مغز، نیمکره های مغزی و ساختار آنها انجام می شود.
پیوند حسی تا حدی توسط سلول های تک قطبی نشان داده می شود که دارای یک آکسون آوران یا آوران هستند و به عقده های نخاعی یا جمجمه ای تعلق دارند. و همچنین گره های اعصاب واگ که ساختاری شبیه به سلول های نخاعی دارند. این پیوند شامل سلول های دوگل نوع II است که اجزای گانگلیون های اتونومیک هستند.
لینک درج
پیوند میانی در سیستم عصبی خودمختار برای انتقال از طریق مراکز عصبی تحتانی، که عقده های اتونوم هستند، خدمت می کند و این کار از طریق سیناپس ها انجام می شود. در شاخ های جانبی نخاع قرار دارد. ارتباط مستقیمی از پیوند آوران به نورون های پیش گانگلیونی برای ارتباط آنها وجود ندارد؛ کوتاه ترین مسیر از نورون آوران به نورون انجمنی و از آن به نورون پیش گانگلیونی وجود دارد. انتقال سیگنال به و از نورون های آوران در مراکز مختلف با تعداد متفاوتی از نورون های داخلی انجام می شود.

به عنوان مثال، در قوس رفلکس اتونوم نخاعی، سه سیناپس بین واحد حسی و عاملی وجود دارد که دو سیناپس در داخل و یکی در گره اتونوم قرار دارند که نورون وابران در آن قرار دارد.
لینک وابران
پیوند وابران توسط نورون های اثرگذار که در گره های رویشی قرار دارند نشان داده می شود. آکسون های آنها فیبرهای غیر میلینی را تشکیل می دهند که همراه با رشته های عصبی مخلوط، اندام های داخلی را عصب دهی می کنند.
قوس ها در شاخ های جانبی قرار دارند.
ساختار گانگلیون عصبی
گانگلیون مجموعه ای از سلول های عصبی است که شبیه به ضخامت 10 میلی متر هستند. با توجه به ساختار آن، گانگلیون اتونوم در بالا با یک کپسول بافت همبند پوشیده شده است، که یک استروما از بافت همبند شل در داخل اندام ها را تشکیل می دهد. نورون های چند قطبی که از یک هسته گرد و هسته بزرگ ساخته شده اند، از یک نورون وابران و چندین نورون آوران واگرا تشکیل شده اند. این سلول ها از نوع سلول های مغزی هستند و سلول های حرکتی هستند. آنها توسط یک غشای شل احاطه شده اند - گلیا گوشته، که یک محیط ثابت برای بافت عصبی ایجاد می کند و عملکرد کامل سلول های عصبی را تضمین می کند.
گانگلیون اتونوم دارای آرایش پراکنده ای از سلول های عصبی و بسیاری از فرآیندها، دندریت ها و آکسون ها است.

گانگلیون ستون فقرات دارای سلول های عصبی است که به صورت گروهی چیده شده اند و آرایش آنها دارای نظم مشخصی است.
عقده های عصبی اتونومیک به دو دسته تقسیم می شوند:
- نورون های حسی که نزدیک به ستون فقرات یا ناحیه مرکزی مغز قرار دارند. نورون های تک قطبی که این گانگلیون را تشکیل می دهند یک فرآیند آوران یا آوران را نشان می دهند. آنها برای انتقال آوران تکانه ها خدمت می کنند و نورون های آنها هنگام انشعاب فرآیندها یک انشعاب تشکیل می دهند. این فرآیندها اطلاعات را از محیطی به نورون آوران مرکزی - این فرآیند محیطی و مرکزی است - از بدنه نورون به مرکز مغز منتقل می کند.
- از نورون های وابران تشکیل شده است و بسته به موقعیت آنها پاراورتبرال، پیش مهره ای نامیده می شوند.
عقده های سمپاتیک
زنجیره های پاراورتبرال گانگلیون در امتداد ستون فقرات در تنه های سمپاتیک قرار دارند که در یک خط طولانی از قاعده جمجمه تا دنبالچه امتداد دارند.

شبکه های عصبی پیش مهره ای نزدیک به اندام های داخلی قرار دارند و محل قرارگیری آنها در جلوی آئورت متمرکز است. آنها شبکه شکمی را تشکیل می دهند که از شبکه های مزانتریک خورشیدی، تحتانی و فوقانی تشکیل شده است. آنها توسط نورون های آدرنرژیک حرکتی و کولینرژیک مهاری نشان داده می شوند. همچنین ارتباط بین نورون ها توسط نورون های پیش گانگلیونی و پس گانگلیونی انجام می شود که از واسطه های استیل کولین و نوراپی نفرین استفاده می کنند.
عقده های داخل دیواره دارای سه نوع نورون هستند. توصیف آنها توسط دانشمند روسی A.S. Dogel انجام شده است که هنگام مطالعه بافت شناسی نورون های سیستم عصبی خودمختار، نورون هایی مانند سلول های وابران بلند آکسونال نوع اول، سلول های آوران متساوی الاضلاع نوع دوم و سلول های انجمنی را شناسایی کرد. نوع سوم
گیرنده های گانگلیونی
نورون های آوران عملکرد بسیار تخصصی دارند و نقش آنها درک محرک ها است. گیرنده های مکانیکی (پاسخ به کشش یا فشار)، گیرنده های نوری، گیرنده های حرارتی، گیرنده های شیمیایی (مسئول واکنش های بدن، پیوندهای شیمیایی)، گیرنده های درد (پاسخ بدن به محرک های دردناک - آسیب به پوست، و غیره) هستند.

در تنه های سمپاتیک، این گیرنده ها اطلاعات را از طریق یک قوس بازتابی به سیستم عصبی مرکزی منتقل می کنند، که به عنوان سیگنالی در مورد آسیب یا اختلال در بدن و همچنین عملکرد طبیعی آن عمل می کند.
وظایف عقده ها
هر گانگلیون مکان، منبع خون خاص خود را دارد و عملکرد آن با این پارامترها تعیین می شود. گانگلیون نخاعی که از هستههای مغز عصببندی میکند، ارتباط مستقیم بین فرآیندهای بدن را از طریق یک قوس بازتابی فراهم میکند. این اجزای ساختاری نخاع غدد و ماهیچه های صاف عضلات اندام های داخلی را عصب دهی می کنند. سیگنال هایی که در امتداد قوس بازتابی می رسند کندتر از سیستم عصبی مرکزی حرکت می کنند و به طور کامل توسط سیستم خودمختار تنظیم می شوند که عملکرد تغذیه ای و وازوموتور نیز دارد.



